In mammals, the visual cells in the retina obtain visual information by perceiving light and then processing it as neural signals. The nerve fibers from the retina run through the optic nerve and the optic chiasm forming the optic tract and are transmitted to the visual relay neurons in the lateral geniculate nucleus (LGN). The axons of the visual relay neurons in the LGN transmit information to the visual cortex through the optic radiations. The genes expressed in the neurons in response to visual stimulation are classified as immediate early genes (IEGs) or late response genes, according to their expression time after stimulation [1]. It is known that IEGs are activated temporarily and rapidly in response to diverse cellular stimulations. Currently, c-Fos, c-myc, and c-jun are the most widely known IEGs. Previous studies have reported that the expression of the c-Fos gene was induced rapidly and temporarily within a few minutes in response to external stimulation [2-4]. Furthermore, c-Fos protein translated by the c-Fos gene is able to induce the transcription factors that can activate or suppress the expressions of specific genes [5]. In the visual transmission system such as the retina, the superior colliculus, and the visual cortex, expression of the c-Fos gene related to light stimulation has been reported [6-8]. Nonetheless, few studies on the expression of the c-Fos gene within the LGN have been performed. Therefore, we examined both the morphological changes and the expression of c-Fos immunoreactivity in the LGN induced by light exposure.
Materials and Methods
For the experiments, albino rats (Sprague-Dawley strain) weighing approximately 250 grams were used. Ten neonatal albino sibling rats were divided immediately after birth into the control group (4 rats) and experimental group (6 rats). In total, five pregnant albino rats (50 neonates) were used in the same way. Therefore, the control groups contained 20 neonates, and the experiment groups contained 30 neonates. In order to manage the albino rats, we adhered strictly to the Guiding Principles in the Care and Use of Animals (US Department of Health, Education and Welfare publication NIH 85-23).
In the control group, each scheduled period (I. immediately after birth, II. 1 week after birth, III. 2 weeks after birth, IV. 4 weeks after birth) contained 5 neonates. As scheduled, regardless of their gender, aspiration anesthesia with ethyl ether was induced between 10 o'clock and 12 o'clock in the morning. The chest was then opened, and the perfusion fixation was performed with 4% paraformaldehyde-0.1% glutaraldehyde solution through the left ventricle of the neonatal albino rats up to 2 weeks after birth and through the ascending aorta of the neonatal albino rats 4 weeks after birth. The brain was then extracted, and post-fixation was performed for 24 hours using the same solution as in the perfusion-fixation.
In the experiment group, immediately after birth, neonatal albino rats were reared in a dark room for the scheduled periods (I. 1 week after birth, II. 2 weeks, III. 4 weeks). Each stage contained 10 neonates. Immediately after the scheduled dark-rearing, regardless of their gender, the rats were exposed to sunlight for 1 hour and anesthetized. The perfusion fixation and the post-fixation were then performed using the same methods as in the control group.
For the preparation of samples for microscope analysis, paraffin sections 7 ┬Ąm thick were made. The samples were washed with acidic alcohol and attached to the glass slides with surfaces that were pretreated with gelatin, and paraffin sections 14 ┬Ąm thick were made and attached to SuperFrost plus glass slides (Fisher Scientific Co., Pittsburgh, PA, USA).
Morphological examination was conducted after deparaffinization using xylene and hydration with different concentrations of alcohol, hematoxylin and eosin staining was performed to examine the general morphological changes of the brain, and luxol fast blue as well as cresyl violet staining (Kluver-Barrera method) were performed to examine the myelinated nerve fibers.
For immunohistochemistry using the c-Fos protein antibody, the prepared samples were treated using the identical method of the deparaffinization and hydration process, washed three times with 0.05 M Tris buffer adjusted to pH 7.6 (Tris buffered saline, TBS), and fixed for 10 minutes using 4% paraformaldehyde-0.1% glutaraldehyde. In order to suppress the activation of endogenous hydrogen peroxide, the samples were immersed in a 3% H2O2-methanol mixture solution for 30 minutes and again washed three times with 0.05 M TBS for 5 minutes. To restore the antigenicity of the c-Fos protein, under the condition that the sample slides were immersed completely in 0.05 M citrate buffer solution and adjusted to pH 7.6, the samples were heated in a microwave at 100Ōäā for 10 minutes and cooled at room temperature. To suppress non-specific reactions, the samples were washed for 15 minutes with 0.05 M TBS-A, 0.5% triton X-100, and 0.05 M TBS-B, where 5% goat serum was added to TBS-A.
As the primary antibody, the anti-c-Fos antibody (rabbit-polyclonal; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) diluted to 1:100 with 0.05 M TBS-B was used, reacted at 4Ōäā for 72 hours, and washed with TBS-A and TBS-B for 15 minutes each. For the secondary antibody, biotinylated goat anti-rabbit IgG (Vector Co., Burlingame, CA, USA) diluted to 1:200 with TBS-B was used, reacted at room temperature for 1 hour, washed with TBS-A and TBS-B for 15 minutes each, and reacted for 1 hour with avidin-biotin-peroxidase complex (ABC method) (Vectastain Elite ABC kit, Vector Co.) at room temperature.
As a dye, 0.33% 3.3-diaminobenzidine tetrahydrochloride (10 mg/40 mL TBS; Sigma, St. Louis, MO, USA) was used. Immediately prior to its use, a 3% H2O2 solution was added, used for staining for 5 minutes, washed with 0.05 M TBS solution and distilled water, and counterstained for one minute using Nuclear Fast Red, before it was then washed with distilled water and mounted with permount after the clarifying process.
All of the stained samples were examined under an Olympus AX-70 light microscope, and pictures were taken using a Leica DC200 digital camera. As a result of the immunohistochemical stain, according to the color intensity in the nucleus of the neurons, reactions were evaluated as negative (-), meagerly positive reaction (┬▒), weakly positive (+), moderately positive (++), or strong positive (+++). Quantitative measurements of the color intensity were performed by 2 independent observers (JHL and TKP) in a masked fashion. For statistical analysis, we scored the results of immunohistochemical staining and used the Mann-Whitney U-test. A p-value of less than 0.05 was considered statistically significant.
Results
The morphological assessment
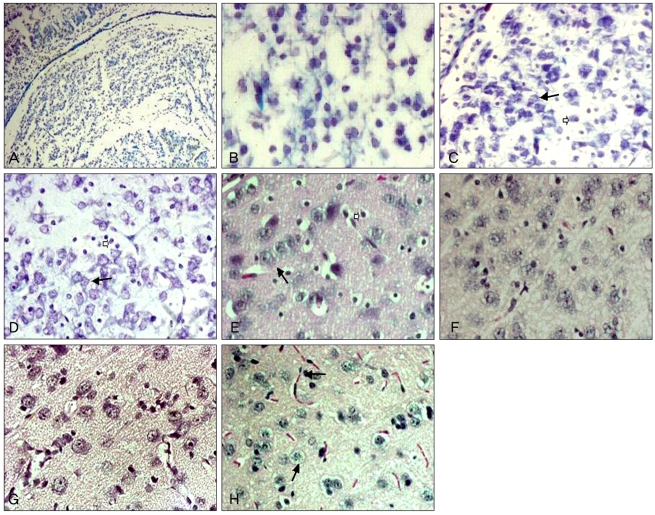
In the control group (immediately after birth), the dorsal and ventral nuclei in the LGN of albino rats could not be distinguished clearly (Fig. 1A). Small undifferentiated neurons with strong basophilic nuclei were observed. The nuclei showing weak basophilic characteristics were surrounded by an extremely small number of neurons in their early differentiation phase, with thin cytoplasm showing strong basophilic characteristics (Fig. 1B). In the control group (1 week after birth), the dorsal and ventral LGN were easy to distinguish based on the primitive intergenicular leaflet, and the myelinated nerve fibers were not developed. In the dorsal LGN, a small number of large neurons with the distinct nucleoplasm and the basophilic cytoplasm were detected. Most neurons contained a meager basophilic vesicular nucleus, and the development of cytoplasm was insufficient; nonetheless, they were clearly distinguishable from astrocytes (Fig. 1C). In the experimental group (1 week after birth), a small number of large neurons with a distinct nucleolus and basophilic cytoplasm were observed in the dorsal LGN. Most neurons had the vesicular nuclei showing basophilic characteristics, and the development of the cytoplasm was meager; nonetheless, they could be clearly distinguished from astrocytes (Fig. 1D). In the control group (2 weeks after birth), large neurons with a vesicular nucleus and abundant cytoplasm and small neurons with a vesicular nucleus surrounded by thin cytoplasm could be clearly distinguished in the dorsal LGN. Astrocytes with a condensed and dark nucleus were observed (Fig. 1E). In the experimental group (2 weeks after birth), more large neurons with many small vesicular structures in the cytoplasm in the dorsal LGN were observed than in the age-matched control group (Fig. 1F). In the control group (4 weeks after birth), large neurons with a vesicular nucleus and abundant cytoplasm and small neurons with a vesicular nucleus surrounded by thin cytoplasm could be clearly distinguished in the dorsal LGN, suggesting normal neuronal development (Fig. 1G). In the experimental group (4 weeks after birth), the cytoplasm of the large neurons became vacuolized. The basophilic characteristics of the Nissl body could not be detected in the dorsal LGN, suggesting neuronal degeneration (Fig. 1H).
The immunohistochemical assessment
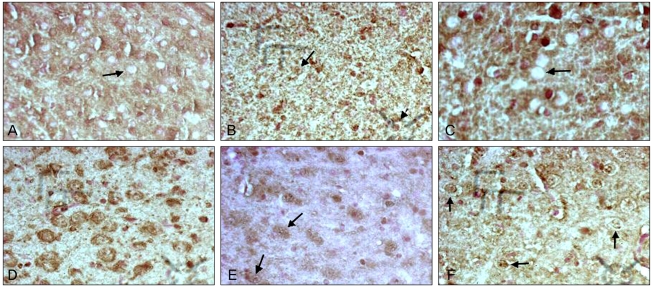
The control group (immediately after birth, 5 neonates) was excluded because there was no comparable age-matched experiment group. In the control group (1 week after birth), c-Fos immunoreactivity was not detected in the nucleus of the neurons in the dorsal LGN (Fig. 2A). In the experimental group (1 week after birth), meager c-Fos immunoreactivity was observed in the nucleus of the neurons in the dorsal LGN (Fig. 2B). In the control group (2 weeks after birth), c-Fos immunoreactivity was not detected in the nucleus of the neurons in the dorsal LGN (Fig. 2C). In the experimental group (2 weeks after birth), granular weak c-Fos immunoreactivity was observed in the nucleus of the neurons in the dorsal LGN (Fig. 2D). In the control group (4 weeks after birth), meager or weak c-Fos immunoreactivity was detected in granular or fibrous patterns in the nucleus of the neurons in the dorsal LGN (Fig. 2E). In the experimental group (4 weeks after birth), moderate or strong c-Fos immunoreactivity was observed in the nucleus of the neurons in the dorsal LGN (Fig. 2F). The color intensity scores in the nucleus of the neurons were 0.00 ┬▒ 0.000 in the control group and 1.00 ┬▒ 0.667 in the experimental group at 1 week after birth (p = 0.013); 1.20 ┬▒ 0.447 in the control group and 2.10 ┬▒ 0.876 in the experimental group at 2 weeks after birth (p = 0.04); and 2.20 ┬▒ 0.447 in the control group and 3.20 ┬▒ 0.789 in the experimental group at 4 weeks after birth (p = 0.04) (Table 1).
Discussion
In mammals, immediately after birth, the LGN and the visual cortex are in the structurally and functionally immature state, and they mature gradually with light stimulation during a certain period after birth, known as the critical period. Generally, the critical period of the visual system in albino rats is from the middle of the second week to about 45 days after birth [9]. Particularly during the critical period, if appropriate visual stimulation is deprived, the visual system will remain in an immature state [10-12].
Together with c-jun, egr-1, and others, attention has been paid to c-Fos as a neurobiological marker of the brain activity related to light stimulation [13]. The c-Fos gene is a type of immediate early proto-oncogene that reacts to mitogenic stimuli, and it has been reported to be involved in diverse cellular processes such as proliferation, differentiation, transformation, and apoptosis. Mammalian c-Fos is a 4 Kb gene with 4 exons, and it contains a 2.2 kb mRNA transcription portion and synthesizes the c-Fos protein composed of 381 amino acids [14]. The Fos family of proteins contains hydrophobic bzip that mediates the protein-protein interactions. The Fos family of proteins forms heterodimers with the diverse Jun family of proteins, thus forming activator protein-1 (AP-1). AP-1 has been reported to play an important role in cellular proliferation, and it was the first discovered mammalian sequence-specific transcription factor. In unstimulated cells, the c-Fos protein is maintained at the basal level, and synthesis is induced temporarily in response to not only biological stimulations such as neurotransmitters and growth factors, but also environmental changes such as light stimulation. In addition, Shaulian and Karin [15] have reported that c-Fos was associated with biological phenomena such as cell cycle progression and tumorigenesis and with increased activity of c-Fos in the skeletal system, the central nervous system, and the hematopoietic system including megakaryocytes during development has been revealed [16-19]. In neurons, the synthesis of c-Fos is induced by neurotransmitters or light stimulation. The axons originating from the neurons in the dorsal LGN are primarily projected to the visual cortex, whereas the axons from neurons in the ventral LGN have been known to be mostly projected to the pretectal area of the midbrain, the superior colliculus, and the structures controlling the reflection related to the visual sense such as the suprachiasmatic nucleus of the hypothalamus.
In the visual cortices of mammals, visual stimulation during the critical period and age-dependent development play important roles in the overall development. Deprivation of visual stimulation due to light deprivation by rearing in the dark or with eyelid sutures impairs the normal development and maturation of various structures in the visual system [12]. This period when the plasticity of neurons is very high is termed the critical period. In albino rats, the critical period of the visual cortex begins at 1 to 15 days after birth, when neonatal albino rats open their eyes, and continues up to 45 days after birth [9]. Mower and Kaplan [8] have reported that the critical period in cats begins approximately 3 weeks after birth, reaching the peak 4 to 5 weeks after birth, subsequently decreasing at 20 weeks, and terminating approximately 1 year after birth [20]. Fosse et al. [13] have reported in a study conducted on cats maintained in a dark environment that appropriate visual stimulation played a more important role in the development and maturation process of the visual cortex during the critical period than does simple age-dependent maturation. Mower and Kaplan [8] have reported that the critical period was prolonged in comparison with that in cats who received normal visual stimulation when cats were raised in a dark room prior to the critical period, whereas the plasticity of the visual cortex was higher in 20-week-old cats reared in a dark environment than it was in the normal controls.
The expression of IEGs is involved in the development and maturation process of the visual system. Ohki et al. [21] have reported that on the 5th day, the 10th day, the 15th day, and the 100th day after birth, albino rats were raised in an environment of alternating 12 hour-unit light/dark cycles and exposed to light for 30 minutes prior to sacrifice. They were sacrificed at 8:30 AM when the c-Fos gene reached the peak level in the inner nuclear layer of the retina or at 2:30 PM when it reached the peak level in the outer nuclear layer of the retina. After examining the expression of the c-Fos gene, it was observed that the expression of c-Fos could not be detected in the retinas of 5-day-old or10-day-old neonatal albino rats; although it was observed in 15-day-old rats. Thus, in neonatal albino rats, the time when the expression of c-Fos began in response to the light exposure is between 11 days and 15 days after birth [21].
In this study, overall c-Fos immunoreactivity in the experiment groups was significantly increased in the dorsal LGN in comparison with that in the age-matched control group. Such results are thought to be attributed to the fact that the albino rats in the control groups received sufficient continuous stimulation by sunlight, thus the normal basal level of c-Fos immunoreactivity was detected. The abnormal morphology of the visual relay neurons suggesting neuronal degeneration was observed in the dorsal LGN in the experimental group. Therefore, if visual stimulations are deprived prior to the critical period, the impairment of the morphological and functional maturation of the LGN will be induced in albino rats, and it is thought that the c-Fos gene plays an important role in the morphological and functional development and maturation in the LGN, similar to other structures of the visual system, such as the retina, the superior colliculus, and the visual cortex.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print